

The fight against cancer continues with “One More”
In the fight against cancer, “one more” matters - one more wedding, one more hug, one more trial. Healthcare professionals won’t stop fighting for their patients, and neither will we.
Learn More at Let’sOutdoCancer.com DetailsWe're Not Waiting for Breakthroughs - We're Working to Create Them
A cancer breakthrough isn’t one moment. It’s decades of research, millions of data points, and resilience in the face of setbacks – all with the hope of challenging what’s possible.
We’re not waiting for breakthroughs – we’re working to create them.
Learn More in USA Today Details
Advancing Cancer Care with Cutting-Edge Science
Powered by science and advanced technologies, we are committed to rapidly advancing novel combinations and next- generation biologics across a wide range of cancers – so that people with cancer can live better and longer lives.
Explore Pfizer Oncology DetailsLatest Articles
Purpose & Ideals
The Mindfulness of Medicine Development: How Scientists Are Driving the Next Wave of Medical Innovations
For millions of patients around the world, the development of a new treatment or vaccine can be life-changing.

Programs & Initiatives
Bridging Global Gaps in Hemophilia Care
Pfizer is proud of its commitment to advancing hemophilia care for more than 40 years, dating back to the introduction of recombinant therapies.

Science & Innovation
The Next Era of Decentralized Clinical Trials: The Clinical Trial Anywhere Model
About Pfizer's initiative to make clinical trials more participant-friendly through the decentralized "Clinical Trial Anywhere" model; a suite of solutions like remote sample collection, alternative study conduct locations, home health, and mobile units to reduce the need for in-person visits.

Products
Every product is the result of 1,500 scientists overseeing more than 500,000 lab tests and over 36 clinical trials before the first prescription.
Pfizer RxPathways
Pfizer RxPathways connects eligible patients to a range of assistance programs that offer insurance support, co-pay help, and medicines for free or at a savings.
Explore RxPathways Details
About
Starting with Charles Pfizer inventing an almond-flavored antiparasite medicine in 1849, our people have always been innovators and trailblazers, committed to finding the next cure.
Learn More About Us Details
Areas of Focus
Revolutionary medicines enable us to enrich and extend life for people living with all types of diseases.

Rare Disease
Explore all of our disease and condition-related resources to learn more about your diagnosis and treatment options.
Gene Therapy
Acromegaly
Duchenne Muscular Dystrophy
Gaucher Disease
Growth Hormone Deficiency
Hemophilia
Sickle Cell
Transthyretin Amyloidosis
Internal Medicine
We’re developing therapies to treat, slow, or prevent disease progression and improve the quality of life for patients with obesity, type 2 diabetes, and cardiovascular and kidney diseases.

Inflammation & Immunology
We look for treatments that provide more than just symptom relief, in order to address the root cause of chronic inflammatory diseases at a molecular level

Vaccines
Vaccines are the single most important innovation in the science of health to significantly reduce the threat of diseases that were once widespread and oftentimes fatal.
Oncology
Cancer treatment needs to be transformed across the entire landscape to significantly improve the lives of cancer patients worldwide.

Anti Infectives
Anti-infectives are medicines that work to prevent or treat infections, including antibacterials, antivirals, antifungals, and antiparasitic medications.
Anti-infectives
Antimicrobial Resistance
AMR Prevention
Antimicrobial Surveillance
Antimicrobial Stewardship
Our Commitment as a Global Leader
Environmental Protection
An Accord for a Healthier World
Where people live shouldn’t impact the quality of their healthcare and income shouldn’t determine health outcomes.
Learn More About The Accord Details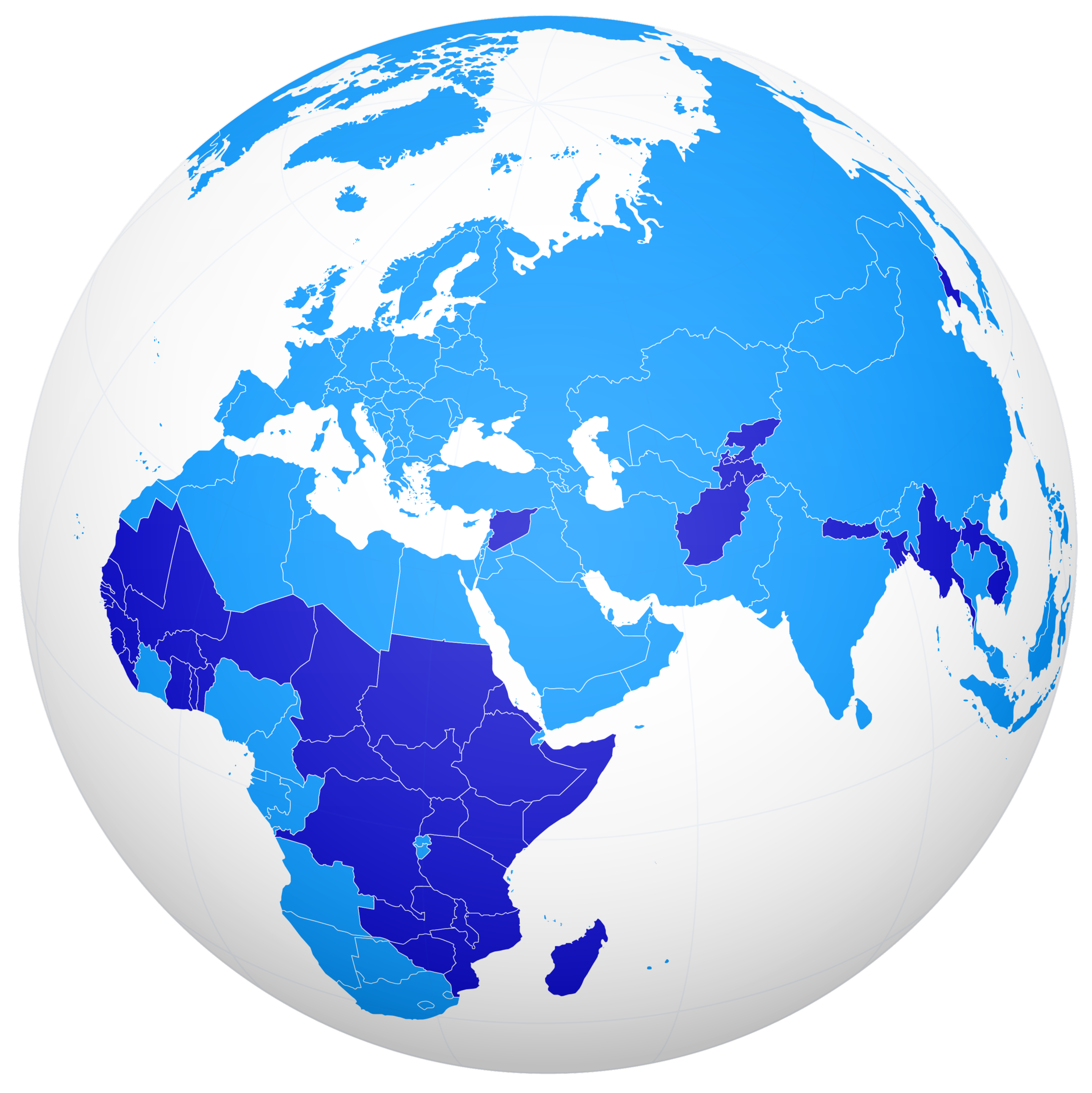

Sign Up for
communications from Pfizer
Receive the latest news from Pfizer in our monthly The Breakthrough newsletter and email alerts on a variety of topics.
Sign up now Details